10 20 30 40 50 60 70 80 MSHHWGYGKH NGPEHWHKDF PIAKGERQSP VDIDTHTAKY DPSLKPLSVS YDQATSLRIL NNGHAFNVEF DDSQDKAVLK 90 100 110 120 125 130 140 150 160 127 GGPLDGTYRL IQFHFHWGSL DGQGSEHTVD KKKYAAELHL VHWNTKYGDF GKAVQQPDGL AVLGIFLKVG SAKPGLQKVV 170 180 190 200 210 220 230 240 DVLDSIKTKG KSADFTNFDP RGLLPESLDY WTYPGSLTTP PLLECVTWIV LKEPISVSSE QVLKFRKLNF NGEGEPEELM 250 260 VDNWRPAQPL KNRQIKASFK CAH2 Human
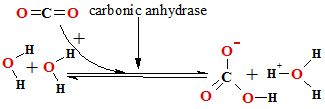
IV. CO2 + 2 H2O<=>H3O++ HCO3- mehanism.
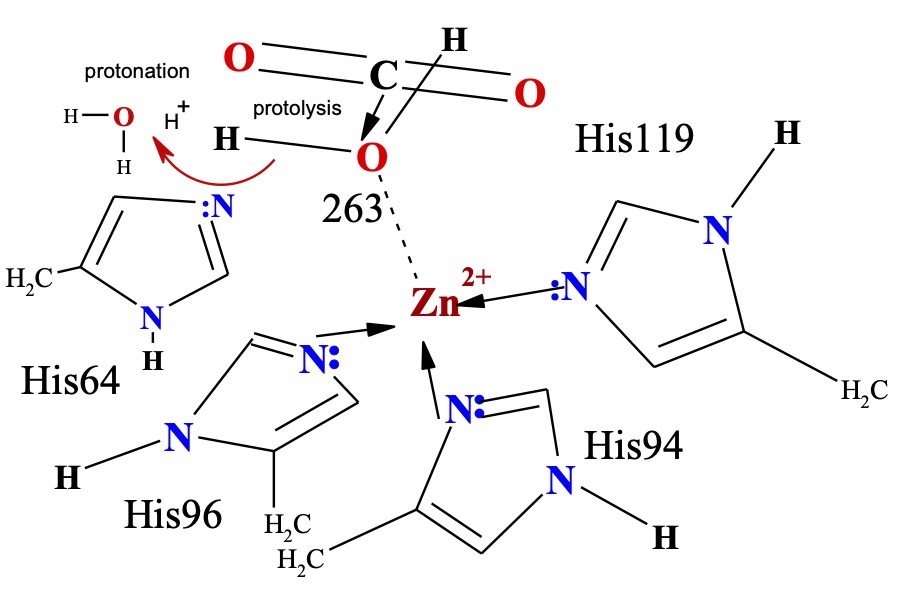
The active site of HCAII. The zinc ion is tetrahedrally coordinated by 3 histidines His-94, His-96, His-119 and catalytic water Wat-263. The proton shuttle His-64, shown in both “in” is linked via Wat-292 at His96 and “out” positions Wat-318 to the catalytic water Wat-263. Hydrogen bonds are depicted as dotted lines, and waters are labeled with numbers only . The deep water Wat-338 sits in a hydrophobic pocketottom of the active site. Wat-318 is in a hydrophilic environment toward the mouth of the active site cone. Zn 2+ CA and CO2 Wat-263 active site 2VVA left at first. Zn 2+ CA and OHCO2- active site 2VVB
References 1.Pharmacol Ther. 1997;74(1):1-20. 1CA2 2.Proteins. 1993 Sep;17(1):93-106. 1CAM 3.J Mol Biol. 1992 Oct 20;227(4):1192-204.Abstract 2CBA 4.J Med Chem. 1995 Jun 23;38(13):2286-91.Abstract 1CNW 5.Martin DP, Hann ZS, Cohen SM. University of California, San Diego , United States. Abstract 4JSW 6.Martin DP, Hann ZS, Cohen SM. University of California, San Diego , United States. Abstract 4JSS 7.PNAS June 30, 2009 vol. 106 no. 26 10609-10613 2VVA=CO2, 2VVB=HCO3- CARBON ANHYDRASE 2CBA 8.January 2004 Molecule of the Month by Shuchismita Dutta and David Goodsell 1CA2 1CAM 1DDZ 1THJ inh 1CNW 9.University Alberta Data Tables Molar Thermodinamic Properties of Pure Substances, http://www.vhem.ualberta.ca/ 10.4.A.M.Suchotina Handbook of ElectroChemistry Petersborg 1981."Chimia"© 11.Daniel C.Harris, "Quantitative chemical analysis". W.H.Freeman and Company ©, 5th ed.1999, p545. 12.J Am Chem Soc.2012 Sep 12;134(36):1472614729. 5G0C 13. Kaksis A. The Biosphere Self-Organization Attractors drive perfect order homeostasis reactions to link bioenergetic with functionally activate oxygen and carbon dioxide molecules. 7th International Conference on New Trends in Chemistry September 25-26, 2021.27-32. Back to the Index...