| Hemoglobin: an O2-transport protein
|
| page 37 |  | | page 38 |
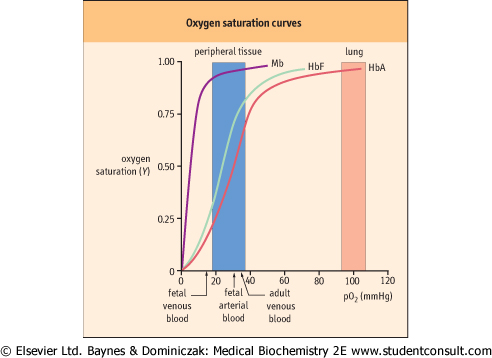
| Figure 4.3 Oxygen saturation curves of myoglobin and hemoglobin. Mb and Hb have different O2 saturation curves. The fractional saturation (Y) of O2-binding sites is plotted against the concentration of O2 [pO2 (mm Hg)]. Curves are shown for Mb, fetal Hb (HbF), and adult Hb (HbA). Also indicated, by arrows and shading, are the normal levels of O2 measured in various adult and fetal blood samples. |
| Hb, the principal O2-transporting protein in blood, is localized in the erythrocytes. As a delivery vehicle, Hb must be able to bind O2 efficiently as it enters the lung alveoli during
respiration and to release O2 to the extracellular environment with similar efficiency as erythrocytes circulate through tissue capillaries. This remarkable duality of function is achieved by cooperative interactions among the globin subunits of Hb.
|
| Quaternary structure of human hemoglobin
|
| Human Hb is a tetramer of two α-globin and two β-globin subunits: α2β2. These subunits are organized in a tetrahedral array, a geometry that predicts several types of subunit-subunit interactions (Fig. 4.4). Experimental analysis of the quaternary structure indicates multiple noncovalent interactions (hydrogen and electrostatic bonds) between each pair of dissimilar subunits, i.e. at the α-β interfaces. In contrast, there are fewer interactions between identical subunits, at the α-α or β-β interfaces. Thus, Hb is more appropriately considered a dimer of heterodimers: (α-β)2. The actual number and nature of contacts differ in the presence or absence of O2 and allosteric effectors. Strong associations within the α-β heterodimer and at the interface between the two heterodimers (Fig. 4.4) are now recognized as major factors determining O2 binding and release.
|
| Interactions of hemoglobin with O2
|
Hb can bind up to four molecules of O2. With its multiple ligand-binding sites, the binding affinity and the fractional saturation of Hb are more complex functions than those of Mb. Consequently, the equation for the fractional O2 saturation curve is modified to
 where n is the Hill coefficient. In a plot of Y versus pO2 when n > 1, the equation for ligand binding describes a sigmoid (S-shaped) curve (Fig. 4.3).
where n is the Hill coefficient. In a plot of Y versus pO2 when n > 1, the equation for ligand binding describes a sigmoid (S-shaped) curve (Fig. 4.3).
|
| Red cell substitutes are allogeneic transfusion alternatives; they are potentially useful during major surgical procedures and in hemorrhagic shock emergencies. The supply-and-demand curves of whole blood and packed red cell availability and use point to an impending crisis and the need to develop alternatives. Three classes of products have been investigated: Hb-based oxygen carriers (HBOC), liposome-encapsulated Hb, and perfluorocarbon emulsions. HBOCs are polymerized forms of hemoglobin; otherwise, the hemoglobin dissociates into monomers (MW ∼16 000 kDa) in plasma and is excreted in urine. Retention of hemoglobin in the kidney may also cause renal failure. Advantages of HBOCs include ease of purification, long-term stability, and minimal immunogenicity. Glutaraldehyde-polymerized bovine Hb (Hemopure®) and diaspirin cross-linked human Hb exhibit increased O2 affinity (P50 = 32-35 mm Hg), diminished cooperativity (n = 1.3-2.1), and decreased sensitivity to allosteric effectors, compared to normal Hb. Both also are more susceptible to autooxidation, a process that leads to significant, although sub-clinical increases in methemoglobin levels. These and other HBOCs have been investigated in clinical trials. In single- and multidose administration, polymerized bovine Hb appears to be well-tolerated in surgical and anemic patients; it has recently been approved for use in some countries. On the other hand, increased mortality with the use of diaspirin cross-linked human Hb has forced the termination of its clinical evaluation. |
| page 38 | | | page 39 |
| Figure 4.4 Model of hemoglobin. Hemoglobin is a tetramer of four globin subunits. Hb is a tetrahedral complex of two identical α-globins (α1 and α2, greens) and two identical β-globins (β1 and β2, purples). With this geometry each globin molecule contacts the other three, creating the subunit interfaces and interactions that define cooperativity. |
| The Hill coefficient, determined experimentally, is a measure of cooperativity among ligand-binding sites, i.e., the extent to which the binding of O2 with one subunit influences the affinity of O2 with other subunits. For fully cooperative binding, n is equal to the number of sites, an indication that binding at one site maximally enhances binding at other sites in the same molecule. In the absence of cooperativity, even with multiple sites, the Hill coefficient would be 1, i.e. binding of one molecule of O2 does not influence the binding of the
next. This is observed for Hb mutants that have lost functional subunit-subunit contacts.
|
| In practice, a Hill coefficient between 1 and n, the theoretical maximum, is commonly observed. The normal value for adult Hb (n = 2.7) is an indication of strongly cooperative ligand binding. Hb has a P50 of 27 ± 2 mmHg, significantly greater than that of Mb. The steepest slope of the saturation curve for Hb lies in a range of pO2 that is found in most tissues (Fig. 4.3). Thus, relatively small changes in pO2 will result in considerably larger changes in the interaction of Hb with O2. Accordingly, slight shifts of the curve in either direction will also dramatically influence O2 affinity.
|
|