| Transition between deoxygenated and oxygenated hemoglobin
|
| As deoxygenated Hb becomes oxygenated, significant structural changes extend throughout the protein molecule. In the heme pocket, as a consequence of O2 coordination to iron and a new orientation of atoms in the heme structure, the proximal histidine and the helix to which it belongs shift their positions (see Fig. 4.1). This subtle conformational change triggers major structural realignments elsewhere within each subunit. In turn, these tertiary structural changes are transmitted, even amplified, in the overall quaternary structure such that one αβ heterodimer rotates ∼15° and slides ∼0.10 nm relative to the other. Because of the inherent asymmetry of the Hb tetramer, these combined motions result in quite dramatic changes within and, more importantly, between the αβ heterodimers.
|
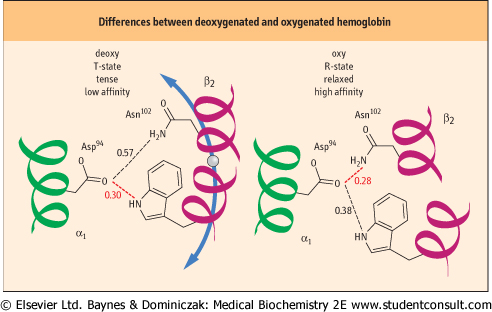
Contact between the two heterodimers (Fig. 4.4) is stabilized by a mixture of hydrogen and electrostatic bonds. Approximately 30 amino acids participate in the noncovalent interactions that characterize the deoxygenated and oxygenated Hb conformations. The two quaternary conformations are known as the T- and R-states, respectively (see Fig. 5.10). In the T-state (tense), interactions between the heterodimers are stronger; in the R-state (relaxed), these noncovalent bonds are, in summation, weaker. O2 affinity is lower for the T-state and higher for the R-state. The transition between these structures is accompanied by the dissolution of existing noncovalent bonds and formation of new ones at the heterodimer interfaces (Fig. 4.5). participate in the noncovalent interactions that characterize the deoxygenated and oxygenated Hb conformations. The two quaternary conformations are known as the T- and R-states, respectively (see Fig. 5.10). In the T-state (tense), interactions between the heterodimers are stronger; in the R-state (relaxed), these noncovalent bonds are, in summation, weaker. O2 affinity is lower for the T-state and higher for the R-state. The transition between these structures is accompanied by the dissolution of existing noncovalent bonds and formation of new ones at the heterodimer interfaces (Fig. 4.5).
|
| page 39 |  | | page 40 |
| Figure 4.5 Noncovalent bonds differ in deoxygenated and oxygenated hemoglobin. In the middle of the interface between the two αβ heterodimers are the residues Asp94 α1 on one heterodimer and Trp37 β2 and Asn102 β2 on the other. Each has side-chain atoms capable of non-covalent interactions. Left: In the deoxygenated T-structure the distance between the Asp and Trp residues favors a hydrogen bond, whereas the distance between Asp and Asn is too great. Right: As a result of the conformational changes that accompany the transition to the oxygenated R-structure, the distance between Asp and Trp is now too large, but that between Asp and Asn is compatible with formation of a new hydrogen bond. Elsewhere along this interface, other bonds are created and broken. An identical alignment of residues and noncovalent interactions is found between the α2 and β1 monomers (Fig. 4.4). Distances are shown in nm. Hydrogen bonds are commonly 0.27-0.31 nm in length. |
| Several models have been developed to describe the transition between the T- and R-states of hemoglobin. At one extreme is a model in which each hemoglobin subunit sequentially responds to O2 binding with a conformational change, thereby permitting hybrid intermediates of the T- and R-states. At the opposite extreme is a model in which all four subunits
switch concertedly; hybrid states are forbidden, and O2 binding shifts the equilibrium between T- and R- states. The molecular structures of deoxygenated and partially and fully liganded Hb have been studied extensively. Despite a wealth of thermodynamic and kinetic data and recent elegant investigations of ligand interactions with Hb crystals and silica-encapsulated Hb, progress toward reconciling inconsistencies with both models has been slow. Much evidence appears to support an allosteric mechanism with features of both models, in part because fundamental questions still remain unanswered. Do the α- and β-subunits differ in ligand affinity? Which subunit binds the first (or releases the last) molecule of O2?
|
|